


Chapitre 2 : Le système visuel humain
Cette description est tirée de [28].
Le système visuel peut être subdivisé en trois sous-ensembles,
représentant le cheminement de l'information visuelle de l'oeil
vers le cerveau. Le couple oeil-rétine, d'abord, officie
comme capteur des signaux lumineux. Ensuite, le nerf optique, le
chiasma
et le corps genouillé latéral permettent la transmission
nerveuse de l'image au
cortex, enfin, où elle est décodée
(figure 2.1). Chaque étape
influe sur le signal transmis. Des pré-traitements sont effectués
mais aussi des pertes y sont occasionnées.

Figure 2.1 : Le système visuel humain
2.1 L'oeil
L'oeil est composé de plusieurs éléments qui ont chacun
une influence dans le cheminement de la lumière et la compréhension
du signal optique par le cerveau1,
(figure 2.2).

Figure 2.2 : L'oeil
Parmi ces éléments, les plus importants sont :
-
la cornée : C'est la couche externe convexe de l'oeil. Elle
est composée de quatre couches successives. Sa courbure dépend
des individus et varie aussi avec l'âge ;
-
l'humeur aqueuse : C'est un liquide salin et alcalin sous pression
qui maintient ainsi la rigidité du globe oculaire ;
-
l'iris, un diaphragme vertical percé en son centre par la
pupille. Il permet, en ajustant la taille et la forme de celle-ci, de réguler
la quantité de lumière atteignant la rétine ;
-
le cristallin, une lentille transparente placée derrière
l'iris. La perte de transparence des cellules cristallines, entraînant
leur mort, s'appelle la cataracte ;
-
le corps vitré, qui constitue les 4/5 du volume de l'oeil.
Composé d'un liquide albumineux sous forme de gelée, il est
parfaitement transparent. Son indice de réfraction varie suivant
la longueur d'onde (d'environ 1.345 pour 400nm à 1.330 pour
700 nm). Ce phénomène s'appelle l'aberration chromatique
et il diminue l'acuité visuelle.
2.2 La rétine
C'est la membrane la plus interne de l'oeil (un prolongement du nerf optique).
Elle reçoit les signaux lumineux, et assure leur transmission au
cerveau par l'intermédiaire du nerf optique. La rétine possède
plusieurs zones où l'acuité visuelle varie.
La fovea est la zone d'acuité maximale. Décalée
de 4 degrés par rapport à l'axe optique, elle contient en
son centre, la foveola où les cellules
photo-réceptrices
sont moins nombreuses. La région périphérique de la
fovea
est colorée en jaune, ce qui accroît l'acuité visuelle
et compense le défaut de cellules photo-réceptrices par rapport
à la fovea. Notons enfin que la zone où émerge
le nerf optique s'appelle la tâche aveugle. Elle ne contient aucune
cellules photo-réceptrices.
Outre une organisation spatiale, la rétine possède aussi
plusieurs couches. La couche externe est composée de cellules photo-sensibles,
les
bâtonnets et les cônes. Ces photo-récepteurs
reçoivent l'information optique par l'intermédiaire de pigments
visuels et doivent la transmettre au cerveau par influx nerveux (figure
2.3)
grâce à plusieurs types de cellules. Les cellules bipolaires,
d'abord, font le lien entre les photo-récepteurs et les cellules
ganglionnaires. Il en existe deux types. Les petites sont associées
aux cônes et les grosses aux bâtonnets. Les cellules horizontales
et amacrines, elles, sont chargées de propager l'information latéralement.
Enfin, l'impulsion visuelle est propagée via les cellules ganglionnaires,
en contact avec les fibres du nerf optique.
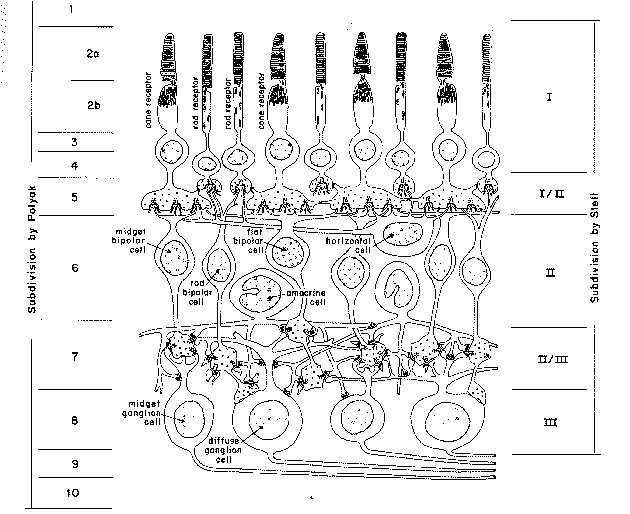
Figure 2.3 : La rétine
Il faut noter qu'il existe trois types de cônes : S, M et L (pour
short,
medium
et long wavelength). Leurs sensibilités maximales sont respectivement
de : 420nm, 530nm et 560nm. Les bâtonnets, eux,
ne sont sensibles que pour des longueurs d'ondes inférieures à
495nm.
Ces photo-récepteurs fonctionnent selon trois modes :
-
photopique le jour, seuls les cônes sont actifs,
-
scotopique la nuit, ce sont les bâtonnets qui fonctionnent,
-
mésopique, où tous les photo-récepteurs sont
actifs.
De plus, leur répartition n'est pas égale dans la rétine.
Ainsi, les cônes ne sont pratiquement présents que dans la
fovea
avec une densité maximale de 150000/mm2.
Celle-ci décroît de manière concentrique. A 40 degrés
de l'axe visuel, on considère la densité des cônes
comme trop faible pour avoir une influence sur la vision.
Les bâtonnets, eux, ne sont pas présents dans la foveola
et très peu dans la fovea. Leur densité maximale est
d'environ 160000/mm2 à 20
degrés. La vision à la périphérie de la rétine
est donc monochromatique et ne sert qu'à la détection des
mouvements. La figure 2.4
illustre cela.
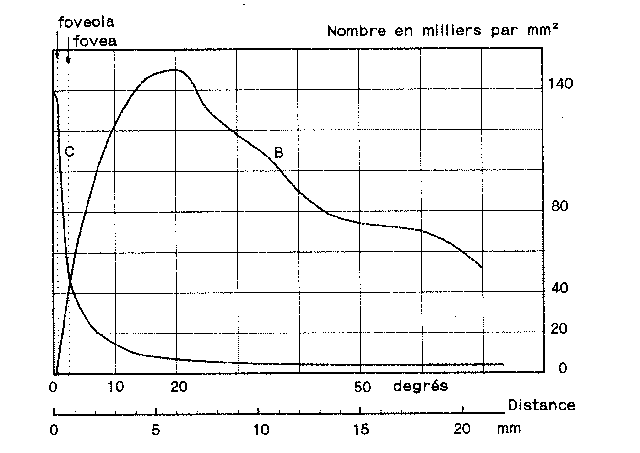
Figure 2.4 : Répartition des cônes et des batonnets
2.3 Transmission de l'information au cerveau
Les deux nerfs optiques se rejoignent au niveau du chiasma. Il y
a alors séparation suivant le champ observé. L'information
gauche est envoyée vers l'hémisphère droit et inversement.
Les reliefs peuvent ainsi être détectés par comparaison
des deux demi-images. Le signal optique, transformé en signal électrique
par la rétine, arrive alors au cortex où il est analysé.
Des phénomènes complexes entrent en jeu. Ainsi, la perception
visuelle dépend des facteurs environnants comme le bruit. L'état
de fatigue ou de stress joue aussi un rôle dans la compréhension
de l'image fournie.
2.4 Les phénomènes importants de la vision
2.4.1 L'accomodation
L'accomodation est la faculté d'adapter l'oeil en fonction de la
distance aux objets observés. C'est le rôle de la cornée
et du cristallin qui, en se déformant, ajustent l'image projetée
sur la rétine. L'hypermétropie et la myopie
sont les troubles visuels issus du dysfonctionnement de l'accomodation.
Ces problèmes sont corrigés par l'ajout d'une lentille convergente
ou divergente devant la cornée.
2.4.2 L'adaptation à la lumière
L'intensité lumineuse à la surface de la terre varie beaucoup.
Ainsi, l'éclairage peut varier de 1015W.cm-2
entre une journée ensoleillée et une nuit sans lune. L'oeil
doit être capable de capter les photons dans l'obscurité mais
aussi de se protéger d'une lumière trop vive ; ceci grâce
à un système d'adaptation à la lumière.
Ce phénomène s'illustre facilement. Lorsque nous quittons
un endroit fortement éclairé pour un autre plus sombre, nous
sommes temporairement aveugles. Après quelques secondes, notre vision
se rétablit. C'est également le cas dans la situation inverse.
Plusieurs modes permettent cela.
La régulation de la quantité de lumière atteignant
la rétine
L'iris est capable de modifier la forme et la taille de la pupille. Une
pupille large laissera passer beaucoup de lumière (mode nocturne),
et inversement. Ce phénomène très rapide n'est cependant
que temporaire. Il permet aux autres dispositifs plus lents de se mettre
en place.
La régulation peut se faire aussi par des mouvements rétiniens.
En effet, en vision nocturne, les bâtonnets sont allongés
et les cônes contractés. L'effet inverse se produit en vision
diurne. Toutefois, ce mécanisme est, à l'heure actuelle,
peut connu.
Le pigment visuel
Composé de rhodopsine2
il a pour charge d'absorber les photons et d'exciter les cellules visuelles.
Cette molécule possède, de plus, la propriété
de blanchir avec l'intensité lumineuse. Son pouvoir absorbant est
alors réduit. Il faut 7 minutes aux pigments visuels des cônes
pour se régénérer et 40 minutes pour ceux contenus
dans les bâtonnets. C'est pour cette raison que nous sommes moins
gênés par une lumière forte soudaine que par l'entrée
dans un tunnel très sombre, par exemple.
Le traitement neural des informations
Ce dernier phénomène est encore mal compris. Cependant, il
semble que les neurones en contact avec les cellules rétiniennes
jouent aussi un rôle dans le contrôle du gain.
2.4.3 L'acuité et la sensibilité
L'acuité est le pouvoir de résolution, c'est à dire
la faculté de déceler de très petits objets. Pour
cela, il faut que l'image fournie par le système optique (la cornée
et le cristallin) soit de bonne qualité. Cela dépend aussi
des caractéristiques physiologiques de chaque rétine. Ainsi,
on compte 7 millions de cônes et 120 millions de bâtonnets
pour seulement un million de fibres dans le nerf optique. L'information
est donc compressée avec pertes. Les signaux reçus par deux
cellules proches seront combinés vers une seule fibre. Plus la distance
entre deux photo-récepteurs telle que deux fibres nerveuses soient
sollicitées est réduite, meilleure est l'acuité visuelle.
Il faut noter aussi que cela dépend de l'intensité lumineuse
des sources observées.
La sensibilité, elle, est l'aptitude à détecter
de faibles quantités de lumière. Elle dépend de la
longueur et de la surface des photo-récepteurs. Ce sont donc les
bâtonnets qui offrent une meilleure sensibilité à la
lumière. C'est pourquoi, il est communément admis que la
région périphérique à la
fovea, plus
riche en bâtonnets, détecte l'information de luminance.
2.4.4 La transduction visuelle
La transduction est la conversion de l'énergie lumineuse en impulsions
électriques. Si les études menées ont porté
sur les bâtonnets, il semble que les cônes fonctionnent de
manière similaire.
La perméabilité des cellules des bâtonnets permet
le passage des ions et provoque une excitation électrique sur le
nerf optique. L'hyperpolarisation des cellules visuelles est régie,
notamment, par l'ion sodium. Cependant, ni le mécanisme en jeu,
ni l'influence d'autres substances telles que l'ion calcium ou le guanosine
3',5' n'ont pu être approfondis pour l'instant.
Les physiologistes ont quand même défini une loi mettant
en relation l'intensité lumineuse et la réponse du récepteur
(cf. figure
2.5). Cette loi prend
la forme d'une sigmoïde.
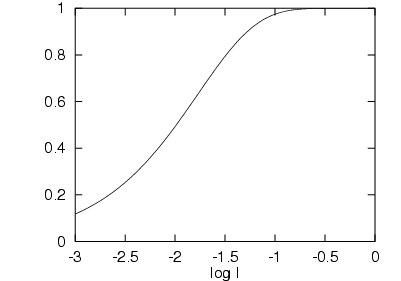
Figure 2.5 : Fonction Transducer
2.4.5 La vision des couleurs
La couleur dépend de la discrimination des longueurs d'ondes dans
le spectre. Deux facteurs permettent de qualifier une couleur : le ton
qui est la longueur d'onde prédominante et la saturation,
c'est à dire la quantité de cette longueur d'onde par rapport
à la distribution spectrale.
Cependant, il convient de faire un distinguo entre la notion de couleur
physique et la perception de celle-ci. Ainsi, certaines espèces
animales ont une vision seulement monochromatique ou dichromatique3
(un seul ou deux types de cônes dans la rétine).
L'homme est considéré comme trichromate. La première
théorie, développée par Young en 1802 puis Helmholtz,
suggérait que la rétine contient trois types de cônes
aux sensibilités spectrales différentes (rouge, vert, bleu).
Certaines critiques furent émises. En 1878, Hering a proposé
une approche alternative. Selon lui, l'homme serait sensible aux différences
de couleurs. Ainsi, les oppositions noir/blanc, rouge/vert et bleu/jaune
décriraient la vision chromatique humaine.
Des études récentes (cf Padgham & Saunders, [26])
semblent attester que les deux théories sont présentes. Les
cônes seraient bien sensibles au rouge, vert et bleu comme indiqué
par Young et Helmholtz. Il y aurait, de plus, un phénomène
chimique dans la rétine. L'information sur les oppositions de couleur
serait aussi transmise au cerveau, le jaune étant obtenu par combinaison
du rouge et du vert.
2.4.6 Le rôle
des fréquences et orientations
De nombreuses études ont montré l'importance de l'organisation
fréquentielle de l'image dans sa compréhension. Pour le cas
des orientations, la littérature est moins abondante.
Fonction de sensibilité au contraste, CSF
Watanabe et al. [32] ont effectué
des mesures sur des sujets humains. Le but était de connaître
leur sensibilité au contraste en fonction de la fréquence
spatiale. La figure 2.6 montre les
résultats.
On observe que les personnes testées sont plus particulièrement
sensibles à des fréquences spatiales comprises entre 1 et
15 cycles par degrés. De plus, au delà de 50 cycles par degrés,
l'oeil ne détecte plus rien.
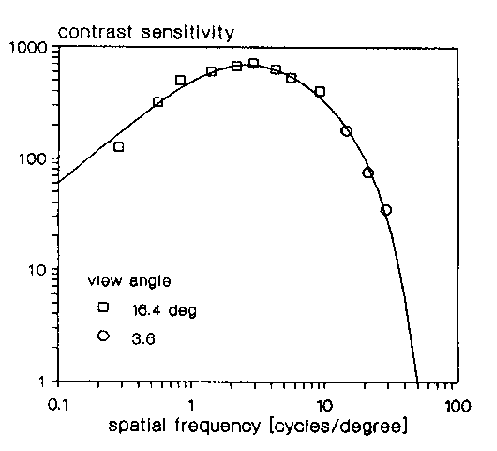
Figure 2.6 : CSF mesurée de Watanabe
Les mesures de Campbell et al. [6]
(figure 2.7) sont similaires.
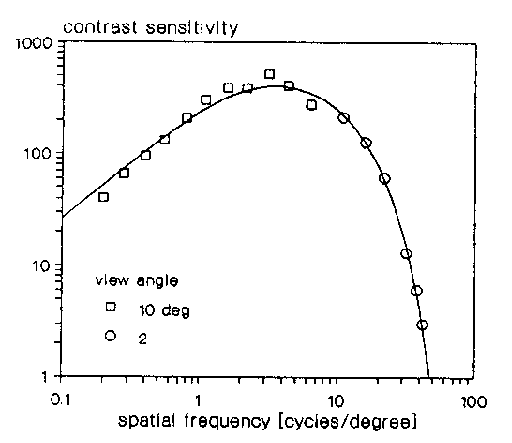
Figure 2.7 : CSF mesurée de Campbell
Des études ont été menées pour trouver une
formule analytique proche des résultats expérimentaux. Parmi
les plus connues, nous pouvons citer la formule de Mannos et Sakrison [19].
Celle-ci est une des premières solutions proposées. Elle
est utilisée dans bon nombre de modèles.
| CSF(f) = 2,6(0,0192+0,114f)e |
|
(2.1) |
Citons aussi la solution de Ngan et al. [24],
plus récente, et donnant de meilleurs résultats.
CSF(f) = (0,31 + 0,69)e-0,29f
(2.2)
Les orientations
Il y a eu très peu d'études menées sur le sujet. Cependant,
il est facile de se rendre compte intuitivement de son influence. Lorsque
l'on regarde une image, nous sommes plus sensibles à l'aliassage
sur des formes verticales ou horizontales qu'obliques. De la même
manière, dans la vie courante, il est plus difficile d'ajuster un
objet avec précision s'il est incliné à 45 degrés.
2.4.7 L'aberration chromatique
Cette sensibilité fréquentielle diffère selon le type
de cônes. Des études ont montré que les cônes
S
ne détectaient rien au delà de 4 cycles par degré,
[22], [27].
Nous devons donc utiliser trois fonctions CSF différentes.
La figure 2.8 montre deux
CSF
achromatique et chromatique.
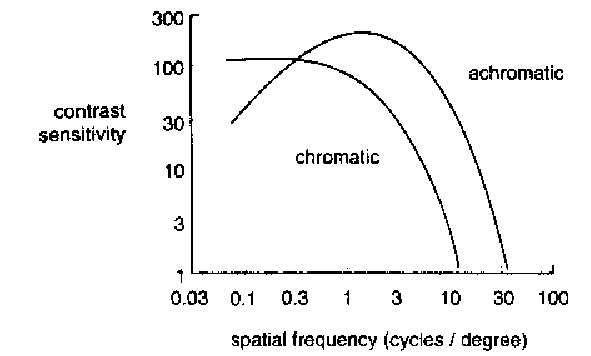
Figure 2.8 : Aberration chromatique
2.4.8 Le masquage
La détection d'une texture n'est pas seulement fonction de la fréquence
spatiale locale. La présence d'autres textures dans la zone influe
aussi. Considérons par exemple une image avec un fond texturé
orienté horizontalement. Un objet ayant la même fréquence
et la même orientation sera difficilement détectable. Si au
contraire, cette objet est orienté verticalement, il sera mieux
repéré. Donc, l'objet devra avoir un poids plus important
en terme d'information significative (figure 2.9).
Figure 2.9 : Phénomène de masquage
2.4.9 La fonction psychométrique
On définit le seuil de détection comme l'inverse de la fonction
CSF.
Si le contraste est inférieur à ce seuil, on considère
que le système visuel ne détecte rien, ou tout au plus une
forme grise peu précise. Au contraire, si le contraste est supérieur,
la zone de l'image est considérée comme significative. Mais,
ce n'est pas une alternative binaire. La transition se fait progressivement.
La probabilité de détection varie donc en fonction de l'écart
du contraste calculé avec le seuil. Lorsque la distance est égale
à 1, la probabilité vaut 75%.
C'est le rôle de la fonction psychométrique. La figure
2.10
montre un exemple. La forme sigmoïde est une bonne simulation du phénomène.
Remarquons aussi la progression linéaire pour des valeurs proches
du seuil.
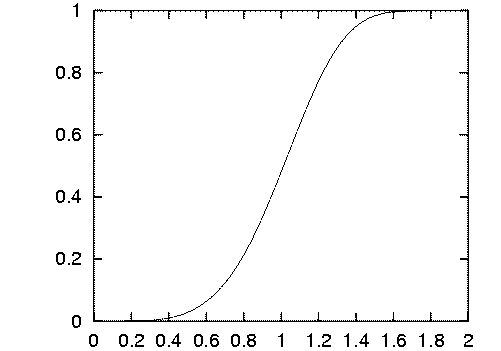
Figure 2.10 : Fonction psychométrique
-
1
-
Le lecteur trouvera de plus amples explications dans [1]
et [29]
-
2
-
pour les bâtonnets. La substance contenue dans les cônes
dépend de leur type mais produit un effet similaire.
-
3
-
Cette caractéristique se retrouve, très rarement, chez
certains individus humains.

